|
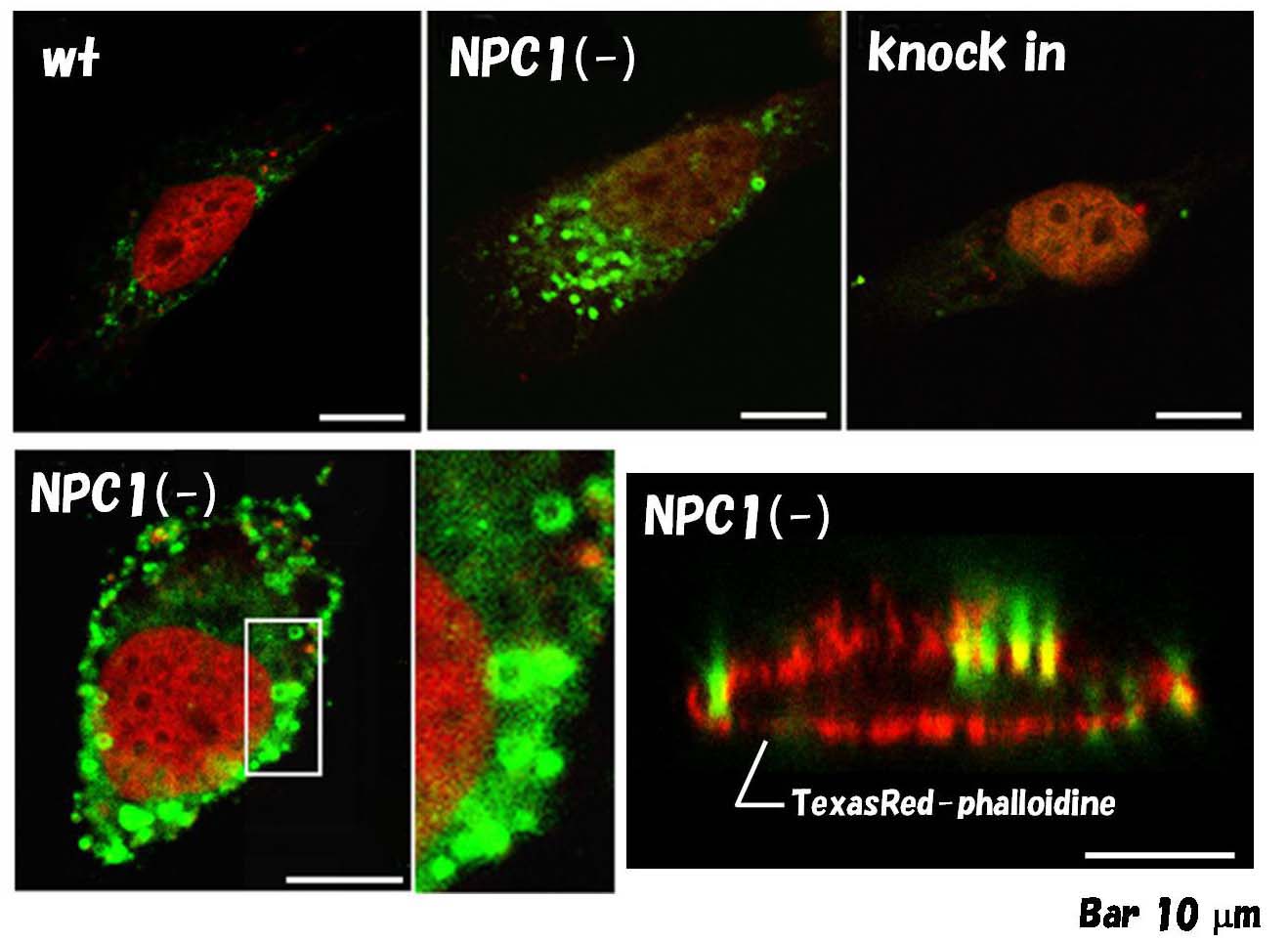
シータ毒素のNPC細胞への結合 |
毒性をなくし、ビオチン化したタンパク質(BCtheta)を用いて、生きている細胞に結合させた。NPC細胞では、細胞表面のベジクルが染まる。
(Ohsaki et al., Histochem. Cell Biol., 2004 [5]) |
| |
|
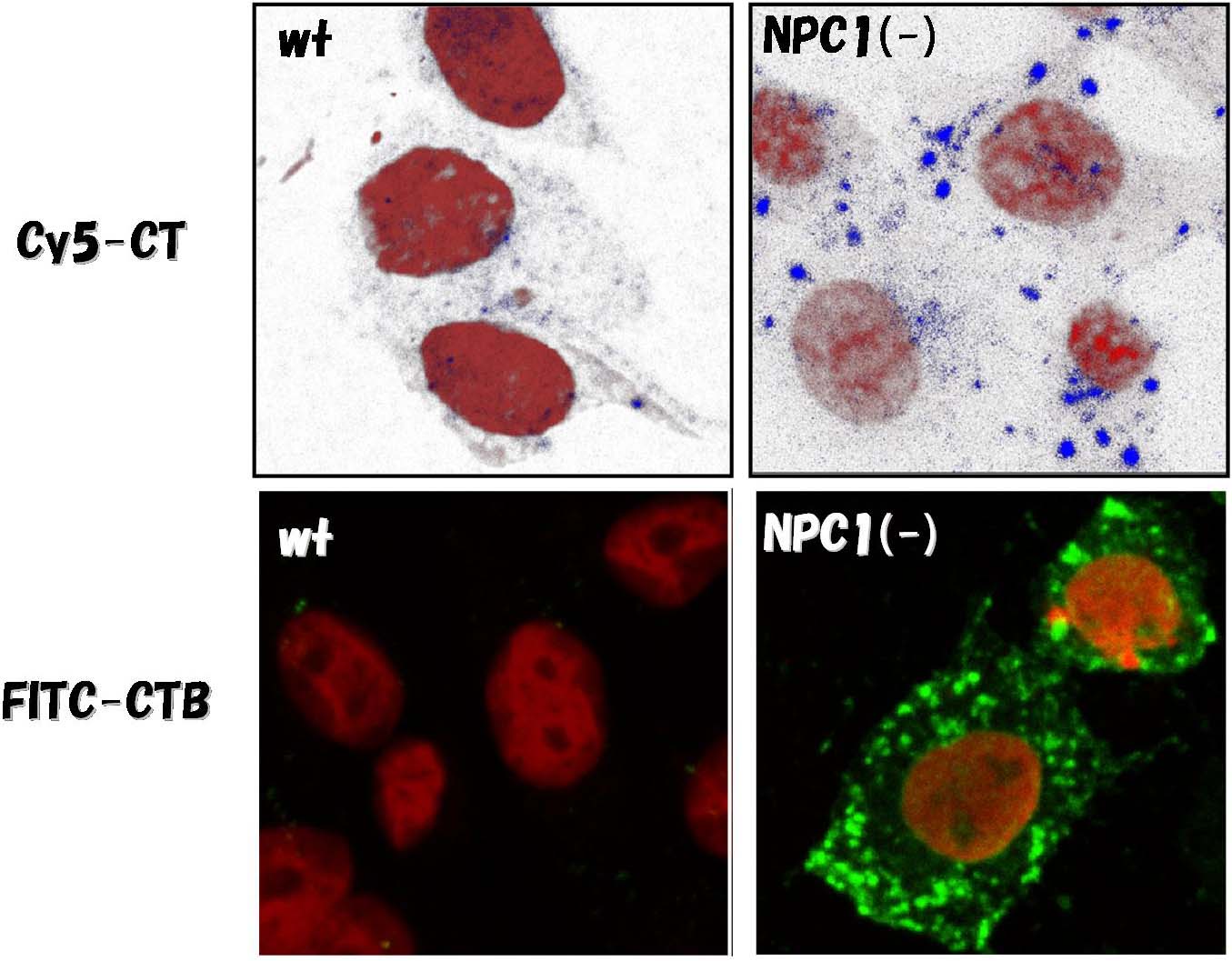
NPC1欠損CHO細胞によるコレラ毒素の取り込み
|
| 上はホロトキシン、下はB subunit。コレラ毒素はGM1に結合する。シータ毒素とは異なり、コレラ毒素は細胞に取り込まれ、early
endosomeにたまる。
(Sugimoto et al., PNAS, 2001 [4]) |
| |
|
|
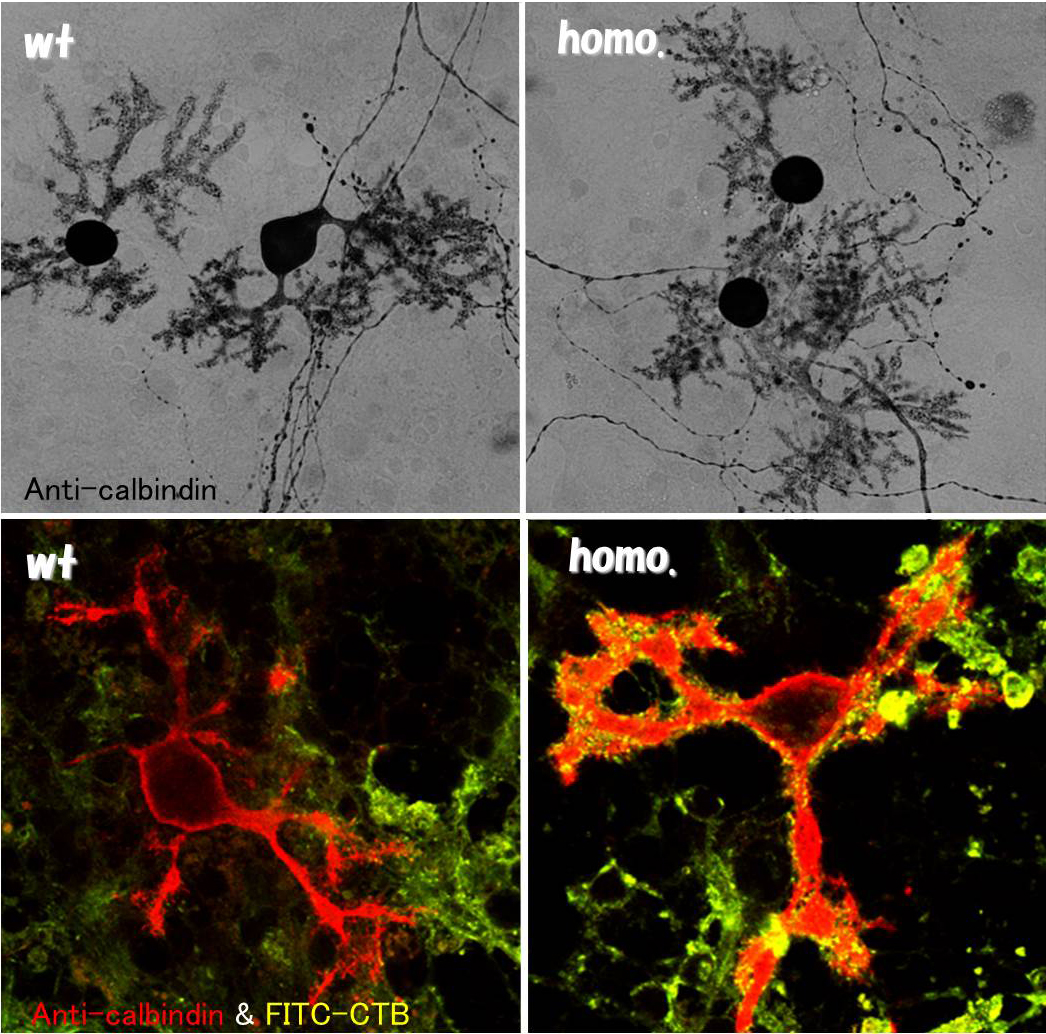
NPCマウス脳からプライマリーカルチャーを作製した。Calbindin染色で形態の変化は認められないが、NPC1欠損細胞では細胞表面および直下がコレラ毒素Bサブユニットで強く染まる。 |
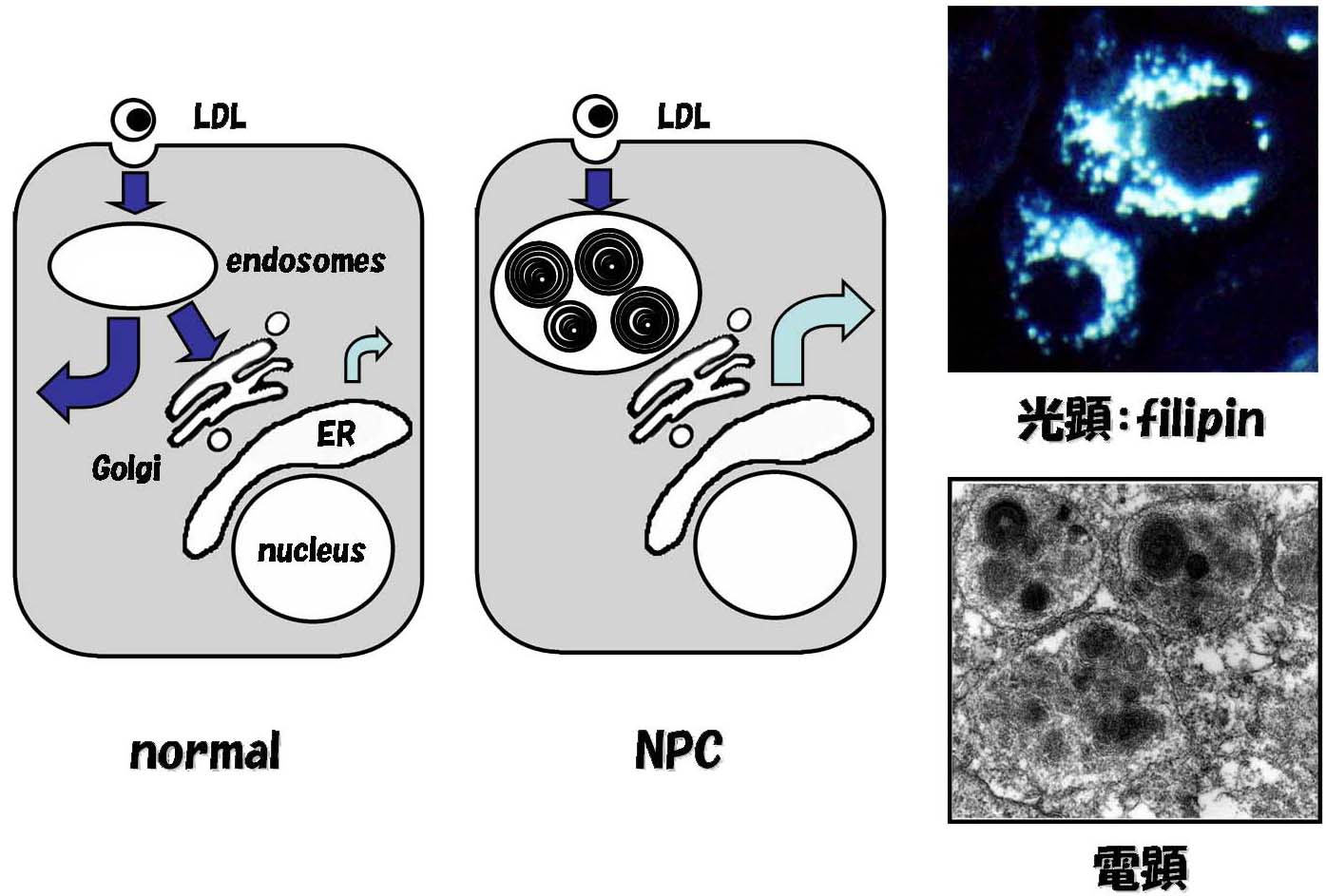
コレステロール輸送に絞って考えると、欠損細胞の表現型からNPC1/NPC2がエンドゾームからの遊離型コレステロールの排出に必要であることは明らかである。細胞内での脂質輸送の機序は小胞輸送とキャリアー蛋白質による輸送に大別されるが、問題はこれらの蛋白質が分子レベルで脂質輸送のどのステップにどのように関与するかということになる。
(1) Neufeld, E.B., Wastney, M.,
Patel, S., Suresh, S., Cooney, A.M., Dwyer, N.K., Roff, C.F., Ohno, K., Morris,
J.A., Carstea, E.D., et al. (1999) The Niemann-Pick C1 protein resides in a
vesicular compartment linked to retrograde transport of multiple lysosomal
cargo. J. Biol. Chem. 274, 9627-9635. [Pubmed]
(2) Higgins, M.E.,
Davies, J.P., Chen, F.W., & Ioannou, Y.A. (1999) Niemann-Pick C1 is a late
endosome-resident protein that transiently associates with lysosomes and the
trans-Golgi network. Mol. Genet. Metab. 68, 1-13. [Pubmed]
(3) Kobayashi,
T., Beuchat, M.H., Lindsay, M., Frias, S., Palmiter, R.D., Sakuraba, H., Parton,
R.G., & Gruenberg, J. (1999) Late endosomal membranes rich in
lysobisphosphatidic acid regulate cholesterol transport. Nat. Cell Biol. 2, 113-118. [Pubmed]
(4) Sugimoto, Y., Ninomiya, H., Ohsaki, Y.,
Higaki, K., Davies, J.P., Ioannou, Y.A. & Ohno, K. (2001) Accumulation of
cholera toxin and GM1 ganglioside in the early endosome of Niemann-Pick
C1-deficient cells. Proc. Natl. Acad. Sci. USA 98, 12391-12396. [Pubmed]
(5) Ohsaki Y., Sugimoto Y., Suzuki M., Kaidoh T., Shimada Y., Ohno-Iwashita
Y., Davies J.P., Ioannou Y.A., Ohno K. and Ninomiya H. (2004) Reduced sensitivity
of Niemann-Pick C1-deficient cells to theta-toxin (perfringolysin O): sequestration
of toxin to raft-enriched membrane vesicles. Histochem. Cell Biol. 121, 263-272. [Pubmed]
last updated Feb. 2009
|